

恐竜図鑑を開くと、必ずと言っていいほど最初の方に登場する、背中に大きな「帆」を持った生き物がいます。それがディメトロドンです。
その見た目から「恐竜の仲間」と思われがちですが、実はディメトロドンは恐竜とはまったく別系統に属する動物であり、むしろ私たち哺乳類に近いグループに属しています。本記事では、ペルム紀の頂点捕食者ディメトロドンの特徴や生態、代表的な種について詳しく解説します。

ディメトロドンは恐竜ではない単弓類

ディメトロドン(Dimetrodon)は、約2億9500万年前から2億7200万年前のペルム紀前期に生息していた大型肉食動物です。
恐竜が登場するのは、その後の三畳紀以降であり、ディメトロドンは恐竜より数千万年も前に繁栄していました。
分類学的にも両者は大きく異なります。
単弓類と双弓類
恐竜は「双弓類」と呼ばれるグループに属します。一方、ディメトロドンは「単弓類」に分類され、これは後に哺乳類へと進化していく系統です。
つまり、ディメトロドンはトカゲや恐竜の仲間ではなく、進化的には私たち哺乳類の遠い祖先側に位置する存在なのです。
単弓類の頭骨には、眼の後方に「側頭窓」と呼ばれる穴が一つだけ存在します。この構造は顎の筋肉を発達させる空間として機能し、強い咬合力を生み出す基盤となりました。
一方、恐竜や爬虫類が属する双弓類では、側頭部に上下二つの穴が開いており、頭骨の軽量化と運動性の向上に適した構造を持っています。単弓類は「噛む力」を重視した進化を選び、双弓類は「動きやすさ」を重視した進化を進めました。この違いは進化の方向性そのものを分け、単弓類はやがて哺乳類へとつながる系統を形成することになります。
最大の特徴「背帆」は何のためにあったのか

ディメトロドン最大の特徴は、背中から突き出した長い骨と、それを覆う皮膚によって形成された巨大な帆です。この構造の役割については、現在も研究が続けられています。
体温調節説
もっとも古くから支持されてきたのが体温調節説です。
帆には多くの血管が通っていたと考えられており、朝は太陽光を受けて体を温め、暑い時は放熱することで体温を調整していた可能性があります。これにより、他の動物より早く活動を開始できたとも考えられます。
ディスプレイ説
近年有力視されているのが、ディスプレイ説です。
帆は異性へのアピールや、ライバルへの威嚇など、視覚的な誇示のために使われていた可能性があります。現生動物でも、派手な飾りを持つ例は多く、この説は比較的現実的と考えられています。
その他の説
過去には、カモフラージュ説や水中推進説なども提唱されましたが、現在ではほとんど支持されていません。
現時点では、体温調節とディスプレイの両方の役割を兼ねていた可能性が高いと考えられています。
「二種類の歯」を持つ進化的に重要な捕食者

ディメトロドンという名前は、「2種類の歯」を意味するギリシャ語に由来します。
実際にディメトロドンの歯は、以下のように分化していました。前方には獲物を突き刺す長い牙状の歯を持ち、後方には肉を切り裂く鋭い歯がありました。
このような歯の分化は、単純な円錐形の歯しか持たない爬虫類型動物とは大きく異なります。
これは、後に哺乳類が持つ門歯・犬歯・臼歯の分化へとつながる重要な進化段階であり、ディメトロドンはその初期形態の一例といえます。
単なる肉食動物ではなく、「咀嚼能力の進化史」においても重要な存在なのです。
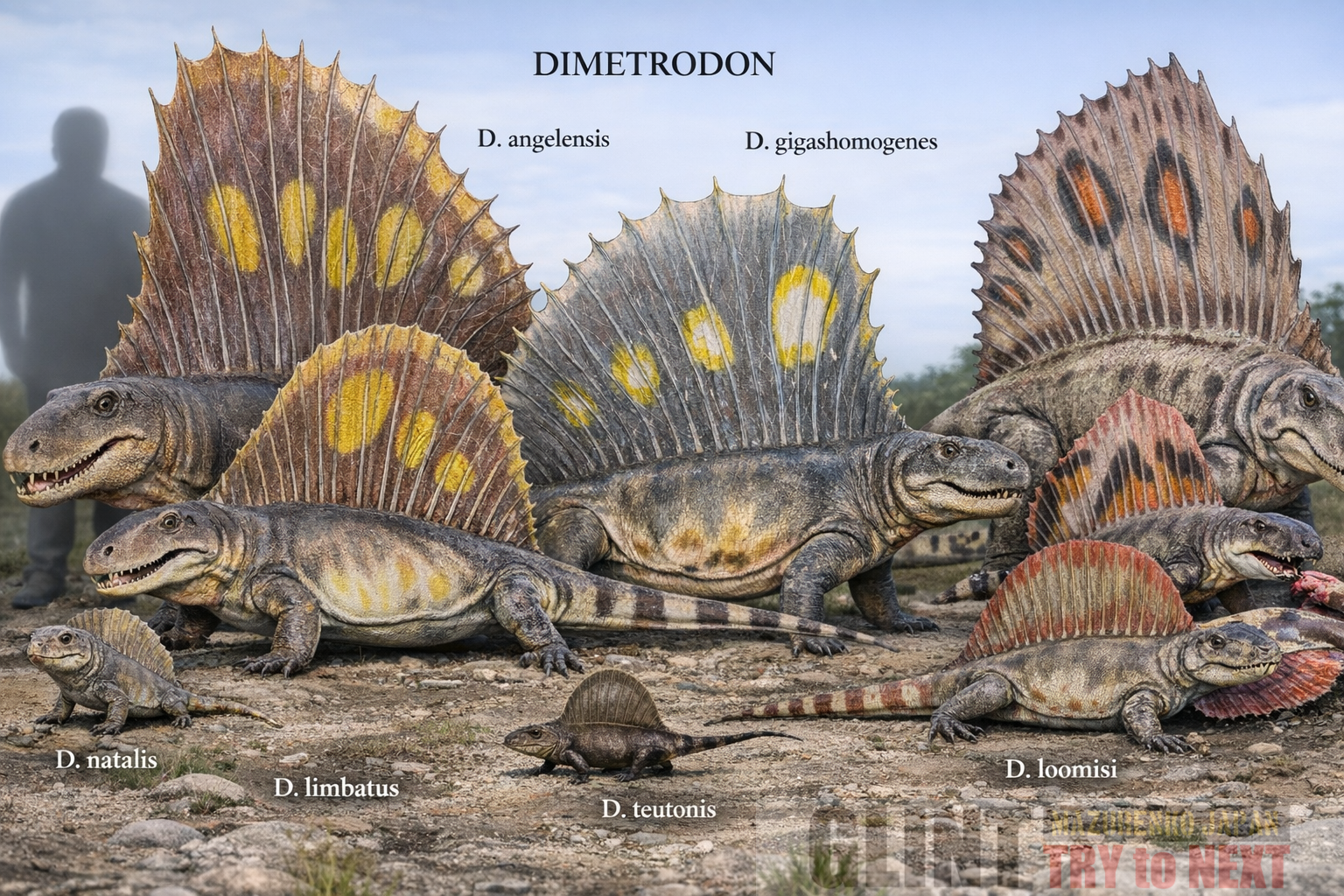
代表的なディメトロドンの種類
現在、ディメトロドン属には10種以上が知られています。その中でも代表的な種を紹介します。
ディメトロドン・グランディス(Dimetrodon grandis)

ディメトロドン属最大級の種です。全長は3.5メートルを超え、当時の生態系における頂点捕食者でした。
主に北米テキサス州周辺から化石が発見されており、巨大な体と発達した帆を持つ姿は、まさにペルム紀の王者と呼べる存在です。
ディメトロドン・リンバタス(Dimetrodon limbatus)

グランディスと並ぶ代表的な大型種です。歯の発達が特に顕著で、大型草食動物を積極的に捕食していたと考えられています。
捕食能力の高さから、当時の食物連鎖の最上位に位置していた可能性があります。
ディメトロドン・ナタリス(Dimetrodon natalis)

全長約1メートル程度の比較的小型種です。
初期的な特徴を多く残しており、ディメトロドンがどのように大型化していったかを研究する上で重要な種とされています。
ディメトロドン・ギガショモゲネス (Dimetrodon gigashomogenes)

全長3メートル以上に達する、属内でも後期に登場した大型種です。最大種である D. grandis に匹敵する体格を持ち、より洗練された頑丈な骨格への進化が見られます。テキサス州のクリアフォーク層から産出しており、当時の過酷な乾燥環境に適応した、ペルム紀における進化の極致とも言えるハンターです。
ディメトロドン・ルーミシ (Dimetrodon loomisi)

全長約2.5メートルほどの中型種で、テキサス州のクリアフォーク層から発見されています。他の大型種に比べて頭骨が低く、細長い形状をしているのが特徴です。その独特な顎の構造から、他の種とは異なる獲物(例えば水辺の小さな獲物など)を好んで食べていた「専門特化型」のハンターであった可能性が指摘されています。
ディメトロドン・ミレリ (Dimetrodon milleri)

全長約1.7メートルほどの比較的小型の種で、テキサス州の初期ペルム紀の地層から発見されています。ディメトロドン属の中でも最も初期に分岐した種の一つと考えられており、大型種に見られるような劇的な特殊化が進む前の、基礎的な骨格形態を留めています。小型ながらも鋭い歯と帆を既に備えており、その後の大型化への進化の起点を知る上で極めて重要な種です。
ディメトロドンが示す哺乳類進化の出発点
ディメトロドンが生きていたペルム紀末には、地球史上最大規模の大量絶滅が起こりました(P-T境界大量絶滅)。
ディメトロドン自体はその前に姿を消しますが、単弓類の系統は生き残り、やがて獣弓類を経て初期哺乳類へと進化していきます。
つまり、ディメトロドンは直接の祖先ではないものの、私たち哺乳類へと続く進化の流れの中に確かに存在していた重要な存在なのです。
怪獣のような外見の裏側には、「哺乳類の夜明け」とも言える進化の萌芽が刻まれていました。ディメトロドンが今も多くの人を惹きつける理由は、そこにあるのかもしれません。
当サイトの人気生物学記事

【生物学のトップページ】
【古代生物種類一覧図鑑】
【恐竜類の種類一覧図鑑】
【絶滅巨大哺乳類の図鑑】
【巨大生物種類一覧図鑑】
【深海魚一覧・種類図鑑】
【超危険生物の画像図鑑】
【世界猛獣種類一覧図鑑】
【世界珍獣一覧種類図鑑】
【有袋類の一覧種類図鑑】
【クジラの比較種類図鑑】
【世界サメ種類一覧図鑑】
【世界奇虫種類一覧図鑑】
【世界のクラゲ種類図鑑】
人気企画「空想生物学」仮想進化研究室

【空想生物学】地球史の「もしも?」を科学的に再構築する仮想進化研究室